Alarm: ez a Y DNA Haplogroupokkal foglalkozó sorozatom úgy keletkezett, hogy miután belefutottam a témába, és számomra teljesen új volt, érdekes játéknak tekintettem, (sok helyütt direkt rájátszással,) és úgy fogtam az "ismertetésébe", hogy magam is közben ismerkedtem vele. Tehát inkább fogható fel szubjektív naplónak az ismerkedésről, mint tényhalmaznak. Természetesen a felkutatott források révén, folyamatosan applikálódott bele egyre több tényként kezelhető kutatási eredmény, -bár minthogy alig több, mint 10 éve született meg az első haplocsoport filogenetikus fa, és a kutatások azóta is folynak, az ún. nevezett szakirodalomban is egymástól jelentősen különböző adatokra bukkanok, főleg az eredet (kor) meghatározások esetében. Saját gondolatok, elképzelések, feltételezések keverednek itt mások (sokszor enyémnél is "őrültebb") hipotéziseivel, tényekként kezelhető ismeretanyaggal. Bár a fejezetek nyilván elkezdődnek és véget érnek, mégsem lezártak, ha valamit érdemesnek találok rá, hogy utólag csatoljam a már leírtakhoz, hasonlóan ehhez a megjegyzéshez, zöld színnel fogom feltüntetni. Az egész sorozat csak egy gondolatébresztő kísérlet, szándéka szerint. Ezt azért is tartom szükségesnek előrebocsátani, mert magyarul olyan kevés anyag van még erről, hogy a google kereső mindjárt a második oldalon kidobta nekem saját kis irományomat, DNS haplocsoport címszó alatt. Ez kemény :)
Tehát fogja fel úgy mindenki, tudományos
mesesorozatot olvas, minden konklúzió nélkül!

Miért, hogy a föl-fölemelkedett Derék Boszorkányt
mindig visszarántja Valaki,
kit Tudósnak is neveznek, Kutatónak is, Genetikusnak is, Professzornak is?
kit Tudósnak is neveznek, Kutatónak is, Genetikusnak is, Professzornak is?
A Derék Banyaember már-már készül
felvilágosítani az emberiséget, de Perazim hegyén fölkél az ...Úr.
Egyik bejegyzésemben megemlítettem, hogy magyarul
alig találtam valamit.
Csak azt nem tettem hozzá, akkortájt még nem is
nagyon kerestem.
Ám kéváncsiság kicsiny férge csak addigelé
rágott agyamban, hogy neki eredtem egy alaposabb rákeresésnek, ahelyett, hogy a
beígért Mindent Elsöprő Összegzésnek láttam volna neki, ígéretemhez egyébként
is híven.
Sejtettem én, hogy előzőleg nem mondhattam teljesen
valóságosat, hiszen már 2003-ban felállították azt a filogenetikus fát,
azóta csak kell, hogy legyen hazai irodalma a neten....
És mibe futottam?
Először egyenest egy Péhádés értekezésbe,
bizonyos Kovácsné Csányi Bernadett követte el ellenem a merényletet.
"A honfoglalás kori, valamint magyar és székely
populációk apai ági genetikai kapcsolatrendszerének vizsgálata,"
ez értekezésének a címe.
 | |
Az még csak nem nem volt baj, hisz az eredmény dettó
az eredményt hozta, amit én az egyik fejezetben megjövendöltem :),
(A székelyek sokkal inkább hordozzák.
Közelebb vannak a Fekete -Tengerhez, a
Kaukázushoz, a Balkánhoz, több bennük az indián-fajta "török" vonal
is. Romániában több az örmény, a cigány, a zsidó is. 7.fejezet)
Nagyobb probléma, hogy felvezetésképpen mindjárt az
enyémtől eltérő kronológiát adott meg, holott én azt is egy professzőr
úrtól kukkoltam le.
Továbbá találtam egy hason témájú prezentációt
is, amely szintén a magyarok genetikai
összetételével foglalkozik :), ezt, bizonyos Dr. Pamzsev
Horolma hozta össze :)
Indítsunk Dr. Horolmával, ez azért is üdvös lesz,
mert egyben átismételhetjük vele az előző részben tanultakat :)
És most nézzük, hogyan adja ugyanezt elő Bernadett:
:)
"A két legrégebbi haplocsoport, az A és a B
klaszter, melyeknél az
M168 marker vad típusú allélja fordul elő, szinte
kizárólag csak Afrikában van jelen.
A kontinensen széles körben elterjedtek, de a
gyakoriságuk alacsony (Underhill et al. 2000).
Underhill és munkatársai (2000) 44 000
évben határozták meg
az M168 marker korát, mely az
Afrikából történő kivándorlást jelzi. Thomson és munkacsoportja
(2000) hasonló dátumot kapott a marker korára vonatkozóan: 40 000 év (95%
konfidencia intervallum: 31 000-79 000 év).
Az M168 marker derivált allélját
hordozó Ősünket nevezhetnénk Eurázsiai Ádámnak is, hiszen ő minden ma
élő, nem afrikai férfi utolsó közös őse valószínűleg észak-kelet-Afrikában,
a jelenlegi Szudán, vagy Etiópia területén élt.
A fiai és unokái által véghezvitt utazás határozta
meg az emberi
történelem elkövetkező lépéseit (Wells 2003).
Leszármazottai 3 nagyobb csoportba különülnek
el.
1. Az M130 marker által jellemzett
Y-kromoszómális vonal
(C haplocsoport)
leszármazottainak jelenkori elterjedése az afrikai
kivándorlás part menti útvonalának hipotézisét bizonyítja. E szerint őseink egy
része körülbelül 50 000 évvel ezelőtt hagyta el az afrikai
kontinenst és indult el Ázsia déli partjai mentén, India déli csücskét érintve,
Délkelet-Ázsiába, majd tovább Ausztrália felé Ennek a vándorlásnak a
régészeti nyomai a tengerszint emelkedése következtében ma nagyrészt rejtve
vannak (ötvenezer éve mintegy 100 méterrel alacsonyabb volt a
tengerszint).
M130-as kromoszómát nem találtak Afrikában, ami azt
jelzi, hogy az M168-ból valahol Ausztrália felé, útközben keletkezhetett.
Az M130-as marker ugyanakkor a kelet-ázsiai
populációkban, köztük a kínaiakban is megtalálható, illetve magas gyakoriságban
van jelen Észak-kelet Ázsiában, főképp Mongóliában és
Szibériában, jelezve, hogy jelentős északi irányú expanzió is
lezajlott Délkelet-Ázsia felül
E mellett a genetikai adatok azt igazolják,
hogy az M130 vonal az elmúlt 10 000 év során elterjedt
Amerikában is. A rendelkezésre álló adatok a partvidéki
vándorlás folytonosságára utalnak, mely szerint 50 000
évvel ezelőtt elindult Afrikából, kelet felé haladva elérte
Indiát, Délkelet-Ázsiát, végül Ausztráliát, majd észak felé fordulva a
Sarkvidék és Amerika felé vette az irányt. A partvidéki marker ősrégi
viszonyt tár fel a kiterjedt térségek lakói közt (Wells 2003).
2. A második csoportot az M89 marker jelöli (az F
szuperhaplocsoportot jellemzi), amely meghatározza a nem
afrikaiak fő Y kromoszóma vonalát.
M89-es Y kromoszómák nincsenek sem Ausztráliában,
(az őslakosok körében!), sem Délkelet- Ázsiában, de elég magas
gyakorisággal fordulnak elő Északkelet-Afrikában.
Ez alapján arra következtethetünk, hogy az M89 kicsivel később alakult
ki (körülbelül 45 000 éves), mint az M130, egy olyan
populációban, amely hátramaradt Afrikában azután, hogy egy csapat a
tengerpartot követve elindult Ausztrália felé. Ezek az M130 nélküli emberek kolonizálták elsőként a
Közel-Keletet. Az M89 marker északkelet-afrikai és levantei jelenléte,
valamint Levante felső paleolitikus lelőhelyeinek kora együtt azt
bizonyítják, hogy a második kivándorlási hullám a Közel-Kelet irányába
zajlott le (Wells 2003). Az Y kromoszóma tanúsága szerint nagyon
kevés ma élő európai tudja genetikai eredetét közvetlenül a 45 ezer
évvel ezelőtti Levantéig visszavezetni. ők lehetnek azoknak a kis
létszámú közel-keleti bevándorlóknak a leszármazottai,
akik elsőként hoztak
be felső paleolit markereket Európába.
Az M89 leszármazási vonalon számos különbözı
haplocsoport (G-R) alakult ki az azokat meghatározó mutációk/markerek megjelenésével.
Az M89 marker által meghatározott
leszármazási vonal képviselői között körülbelül
40 000 évvel ezelőtt valahol a mai Irán
vagy Közép-Ázsia déli területein tűnt fel
az M9 marker, mely a K haplocsoport jellemzője. A
markert hordozó populációkat együttesen eurázsiai klánnak hívjuk.
Az eurázsiai vonalon hozzávetıleg 35 ezer éve jelent
meg Közép-Ázsiában az M45 marker (P haplocsoport), mely két másik, nagyon
fontos vándorlási útvonalhoz vezetett.
Az egyik az Európa irányába visszakanyarodó M173-as vonal (R1
haplocsoport),
a másik az M242 (Q haplocsoport) és M3
(Q3 haplocsoport) markerek által jellemzett útvonal Szibérián keresztül
Amerikába
A felső paleolitikumi anatómiailag modern
európaiak részben Közép-Ázsiából (M173 marker), részben a
Közel-Kelet területéről (M170 marker) eredeztethetık.
A Közel-Kelet területéről a neolitikum
idején is történt egy jelentős bevándorlás
(M172, M201, M35), amely hatással volt az európai génállományra.
3. A harmadik csoportot részben az Y
Alu Polimorfizmus vagy M1 néven ismert marker jelöli,
körülbelül 50 000 évvel ezelőtt alakult ki Afrikában.
Afrikán kívül ez a vonal kettéhasad és lényegében
ugyanazt az útvonalat járja be, mint a másik kettő (Wells
2003).
Ez a polimorfizmus
filogenetikailag egyenértékű az M145 és az M203 markerekkel, mind
a három marker jelen van
a D haplocsoportban (M174 marker jellemzi)
és
az E (M96, SRY4064, P29 markerek
határozzák meg) haplocsoportba tartozó Y kromoszómákon, azok közös eredetét
jelzik.
míg az E haplocsoport elsősorban Afrikában elterjedt, de
származéka, az E3b-M35 klád Európában is jelen van.
Európa paleolitikumi és neolitikumi leszármazási
vonalai Semino és munkatársai közleménye alapján (2000a) az Európában élő
férfiak többsége, több, mint 95%-uk besorolható 10 Y-kromoszómális leszármazási
vonal valamelyikébe.
Munkájuk során 25 európai és közel-keleti helység
1007 férfi lakójának Y kromoszómáján 22 bináris markert vizsgáltak
meg, elsősorban a földművelés terjedésének bizonyítéka után
kutatva. Eredményeik szerint a neolitikus közel-keleti markerek által
meghatározott leszármazási vonalak a modern európaiak kisebb részében vannak
csak jelen.
Következtetéseik szerint, melyek összhangban vannak a mitokondriális
DNS analíziséből származó eredményekkel (Richards et al. 1998,
Torroni et al. 1998), az európai férfiak Y-kromoszómális génkészletének körülbelül
80%-a a paleolitikum idejére vezethető vissza, mintegy 20%-a pedig a
neolitikus közelkeleti bevándorlóktól származik. Semino és
munkatársai szerint az általuk vizsgált markerek földrajzi megoszlása és
becsült kora két paleolitikus és egy neolitikus vándorlási eseményre derít
fényt, melyek hatása kimutatható a modern európai génállományban.
A mai európai férfiak körülbelül 50%-ának Y kromoszómáján van jelen
az M173 mutáció (R1 haplocsoport), mely körülbelül 30 000 éves.
"
Nos Egyelőre Eddig az idézet!
Ha Horolmát eddig összevetjük Bernadettel, azt
kell látnunk, hogy legalább a haplocsortokhoz rendelt mutációk itt még
összecsengenek, nem úgy a melléjük rendelt időpont :)
További levont következtetés, hogy jóval később
szédelegtek ki az Afrikai Ádám fiai,
és hogy három külön hullámban tették, tehát
markánsan elkülönül a C-s csoport, az F-estől, és DE-stől.
Mindezek fényében most úgy látom, hogy az
"afrikai populáció nagy fokú genetikai varianciája" csak azt
jelenti, hogy nincsenek még beleírva a mutációk okozta, determinatív,
öröklődően azonos elváltozások, adott helyeken, lókuszokon nincs meg az a
marker...
De még mindig rohadtul nem értem, az adott mutációk
időpontját milyen elv alapján határozhatják meg, ( a sorrendiséget még
csak-csak, a későbbi hordozza az előbbit is...)
Mint ahogy azt sem értem, ha léteznek
különböző ágak, amelyekből kiesik a többire jellemző egy-egy létrafok, mi
bizonyítja, hogy csak egyetlen közös ős létezhetett? És az is Afrikában?
Mert Darwin 1869-ben ezt írta?
VAn egy kizárólag afrikai halmazunk, az A-, és
B-Haplocsoport, amelyekben annak a bizonyos M168-as markernek még a vad, azaz
módosulatlan allélja fordul elő.
Az összes többi haplocsoportban tehát
jelen van ez a M168-as mutáció, csak az
-ausztráliai, Dél- és Észak-Ázsiai partvidéken
előferdülő C csoportban már ott van az M130-as markeres mutáció is,
- a F-től R-ig terjedőek hordozzák az
alap M168-ason kívül az M89-et, illetve minden
egyes csoport kap még csak rá jellemző újabb mutációkat ezek mellé,
- aztán ott vannak még az Y ALUS (M1-ESEK),
akik az alap M168-on kivül, mind hordják még az M145 és M203-as
mutációkat, illetve még továbbiakat is:
- A D-sek Ázsiában az M174-et,
-Az E-sek Afrikában az M96, SRY4064,
P29 markereket.
|

S ha már egyetlen, közös őst keresünk mindenáron,
nem volna logikus, azt a bizonyos közös, derivált
M168-as markert inkább vadnak tekinteni, és az afrikai A-B-ben előferdülő
"Vadnak" kikiáltott allélt tekinteni mutációnak ?
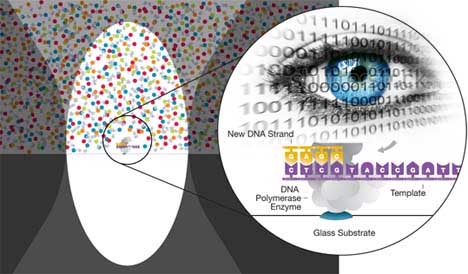
Na jó, leáshatok én akármennyire aprólékos
részletekig, példának okáért sikerült találnom magyar nyelven egy részletes
leírást is, hogyan történik ténylegesen a DNA vizsgálata, a szekvenálás, hogyan
is zajlik a haplocsoport meghatározás...
 |
|
részlet a szekvenálási eljárás leírásából:)
|
dehát, mindez csakis egy
szisztematikusan felépített, aprólékosan kidolgozott rendszer,
bolyghatok benne ítéletnapig, nem fogom felfedezni sosem, ha
az alapkoncepcióban valami bibi van, ha képtelen valaminek a
felfogására, megjelenítésére, ami pedig egyébként jelentős tényezője lenne
rendszernek :(
(Ó én szegény!!
Így sosem fogom tudni bebizonyítani, hogy a
Basic-ADAmunk csak hipotetikus, és valójában a világ
négy sarkán keletkezett annak legalább 4 mutáns variációja, és mikor
ezek megindultak, és mondjuk M130-as C-s Ádám összecsattant az
M89-esekkel, ennek köszönhetően keletkezett egy újabb haplocsoportot generáló
mutáció, nem pusztán a vak véletlen műve a diferenciálódás :(
Brühühü, nem szeretem a vakvéletlent, éljenek a
szerelmi csatározások következtében létre jövő pontmutációk :)
És még jobban éljenek a kivülről eszközölt
génmanipulációk,
és az azt elkövető Annunakik,
avagy az elsüllyedt földrészek szupercivilizált
lényei, akik jól körbemutálták a világot, civilizációjuk elpusztulása után.
Amely haplocsoport(OK) az ő művök, ottan esnek a
csodadolgok.) (tudjuk,a nagyorrúak!!)
Node, most kicsit, (és látszólag) elengedem kedvenc
velőscsontom, és visszakanyarodom, a péhádés remekműhöz:
"... A mai európai férfiak körülbelül
50%-ának Y kromoszómáján van jelen az M173 mutáció (R1 haplocsoport), mely
körülbelül 30 000 éves.
Ez a marker vezet vissza minket Közép-Ázsiába, mely
az egyik kiindulópontja volt Európa modern emberek általi benépesülési
folyamatának (Wells 2003) Semino és munkacsoportja szerint (2000a)
az M173 egy olyan ősi eurázsiai marker,
amit az Európába érkező Homo sapiens vagy magával hozott, vagy
érkezésekor alakulhatott ki.
A marker korát hozzávetőleg 30 000 évben
határozták meg.
Az M173 leszármazási vonal, és a később
ebből egy pontmutáció révén létrejött M17 vonal (R1a1
haplocsoport) Európában egymással ellentétes
földrajzi megoszlást mutat.
Az M17 leszármazási vonal Kelet-Európában
gyakori, (nesze neked "indiság")majd elıfordulása
nyugati irányba csökken.
Az M173 (xM17) vonal (R1b haplocsoport) a legnagyobb gyakorisággal Nyugat-Európában van jelen, ugyanakkor előfordulása keleti irányba csökken. (Egy későbbi vizsgálat (Cruciani et al. 2002) kimutatta, hogy az M173 markert hordozó európai férfiak 2 csoportba oszthatók.
Az egyik csoportba tartozó férfiak Y
kromoszómáján
az M173 marker mellett az M17 marker van jelen,
míg a másik csoportba tartozóknál
az M173 marker az M269 markerrel együtt
fordul elő.)
Semino és munkatársai szerint a két
leszármazási vonal kontinensen belül tapasztalható ellentétes földrajzi
megoszlása úgy értelmezhető,
-mint a
jelenlegi Ukrajna területéről (M17 vonal)
-és az Ibériai félszigetről (M173 (xM17)
vonal)
(mint jégkorszaki „menedékek”-ből
(refúgiumokból) kiinduló elszigetelt népesség-magvak expanziójának bélyege
az utolsó jégkorszak leghidegebb időszakát (Last GlacialMaximum
- LGM) követően.
(A jégkorszak alatt (13-20 ezer évvel ezelőtt)
a különböző humán csoportok arra kényszerültek, hogy az észak-balkáni
menedéket kivéve elhagyják Közép-Európa területét.)
A mai európai Y-kromoszómális génkészlet körülbelül
20%-át jellemzi
az M170-es mutáció jelenléte, mely
az I haplocsoportot határozza meg.
Ez az egyetlen olyan nagyobb Y-kromoszómális
haplocsoport, mely széles körben elterjedt Európában, de gyakorlatilag
máshol nem fordul elő (Semino et al. 2000a, Rootsi et al. 2004).
Ez a mutáció Európában alakulhatott ki a
paleolitikum során (körülbelül 22 000 éve) a Közel-Keletről mintegy
20-25 ezer évvel ezelőtt érkező bevándorlók utódaiban. Az
M170 vonal (I haplocsoport) az M89 marker által meghatározott leszármazási
vonalak (F-R haplocsoport) közé tartozik. (vikingék)
Az európai Y kromoszómák körülbelül 22%-a sorolható be a neolitikus közel-keleti markerek által meghatározott leszármazási vonalak valamelyikébe.
-Az M35 marker (E3b haplocsoport),
-az M172 marker (J2 haplocsoport),
-az M201 marker (G haplocsoport) és az
-M89 marker (F haplocsoport)
által jellemzett leszármazási vonalak előfordulási gyakorisága
a Közel-Kelet felől Európa irányába csökken.
Semino és munkatársai szerint az E3b, a G
és a J2 (valójában az egész J) haplocsoport a neolitikum idején a Közel-Keletről érkező földművelő népek
genetikai hatását tükrözi Európa férfi génállományán.
Vizsgálataik alapján a neolitikus gazdálkodók
genetikai hatása a mediterrán partvidék mentén kifejezettebb. (sasos
görögök, etruszkok, Nagy Sanya)
A fent említett leszármazási vonalakon kívül a 10
Y-kromoszómális leszármazási vonal között szerepelt
a Tat vagy M46 mutáció által meghatározott leszármazási
vonal (N3 haplocsoport) is. Semino és munkacsoportja az általuk vizsgált minták
közül az uráli nyelvű számi, udmurt és mari populációban
detektálta számottevő gyakorisággal a Tat marker jelenlétét.
Ugyanakkor azt találták, hogy a szintén uráli nyelvet
beszélő, magyar populációban egyáltalán nincs jelen ez a polimorfizmus. Zerjal és
munkatársai szerint (1997) a marker körülbelül 4000 éves és egy
Észak-Európára korlátozódó újkeletű uráli migrációra utal..."
Eddig Bernaettünk. Látni való, hogy Ő is lényeges
különbséget tesz, tohonya , Kelet-Európai R1a-sok, és a deli,
kelta, sasorros, R1b-seink között ő, s ami
szintén izgalmas,
rámutat arra, hogy ez az R1a-s társaság ugyan
bizony
sosem járt Indijába, sem nem szkítálkodott a nagy orosz
S elvegyült a Balkáni oduikból előoldalgó
cro-magnoni I-sekkel, meg a Közel-Keletről beszivárgó, ügyi, földmüvelő,
állattenyésztő (és szintén sasorrú)E-s J-s G-s gárdával...
Bernadett kelta R1b-seinket pireneusi
barlangokbó, a nagy kitavaszodáskor előszédelgő csoportosulásnak tartja,
csak azt nem veszi figyelembe, hogy oda mások már elszállásolták az őslakos
I-seket is :), továbbá nem tér ki arra az apróságra, hogy a rákba kerültek
oda?? Mikor egyébként az R csoport valahonnan Belső-Ázsiából ered :)?Emlékszünk
a derék Q-s Indiánusokkal együtt...
Na most kérem szépen, abba az "apróságba"
nem kívánok
beleőrülni ,hogy egész egyszerűen más
markerekkel jellemzi az R1a és R1b-s csoportokra jellemző
mutációt :)
Horolma Az R1b-t M343- ként írja le,
az R1a1 pedig M198-asként jegyzi !! (míg Bernadett
M17-ként)
Mély levegő, s menjünk tovább...
További lényeges különbség, hogy bár egy helyütt
utal ő is az ukrajnai kialakulási területre, másutt már a Kaukázust, és
Iránt emlegeti, nyilván hogy a cuccost be tudja illeszteni, a magát mindmáig
szívósan tartó indoeurópai elméletbe, amit egyébként már sorra tönkre vertek az
azóta megszületett genetikai vizsgálódások eredményei!!!
De erről megint csak később valamikor...
Az is muris, hogy Horolma az R1b-t a cro-magnoni
emberekhez próbálja kötni (tehát nála is szétválik élesen a két R-s gárda!),
dehát a cro-magnoniak már rég tudjuk, hogy valójában a jégkorszakot itt
(Európában) töltő I-sek voltak...(Ezt egyébként utána maga is szépen kifejti és
kibontja :))
Szintúgy stimmel a J-G-E-s közel-keletiekhez
kapcsolása a földművelésnek....
De ami igazán izgi a székely, és az anyaország
között mutatkozó szignifikáns különbség;
- a zsidónak titulált J-s haplocsoport mintegy 17%
százalékos jelenléte,
-a mintegy 6 %-os, a nagymagyarok által méltatlanul
megvetett, halszagú, N csoportos jelenlét (holott az N valahol Ujgúriában ered,
és ragozó nyelvűek, mint pl. a sumérok)
- a titokzatos R1b-s "kelta" csoport
fölénye,
- valamint Q-s "Indián" csoport
megmutatkozása
a székelyeknél... Mindezek után,
vagy a székelyek nem "magyarok",
vagy egyedül ők őrizték meg zárt közösségükben a magyarság valódi jellegét,
akkor viszont szembe kell nézni nagyon erős közel-keleti gyökereinkkel.
A mai magyarok elszlávosodása és elromásodása különben
is hatalmas méretű. ÁM a romáknak még örülhetünk is akár :)
Mert legalább ők hordozzák azt az ősibb változatú
R-es variációt, ami talán valóban köthető (Irán-India) az igazi szkíták egy
csoportjához.
Horrorisztikus videó :)
Külön csemege a székely- populációban erősebb Rb-s
(kelta vonulat,) és a "halszagú" N-es csoport erős jelenléte.
De annak legősibb rétege legalább Észak -Kínában
ered, ott, ahova Körösi Csoma is igyekezett... Még izgalmasabb a P-Q-
indián vonal megmutatkozása!!
És a majom akkor ugrik igazán a vizebe, ha
vizsgálódásainkat kiterjesztjük Ázsiára is, különös tekintettel az R1b-sek
hordozó népeire!
Akkor egyértelműen ki fog derülni, hogy a kurgán
-építős, lovas-nyilas harcmodorú társaság leginkább a ma élő, török
népekhez köthető, az ő fő összetevő csoportjaik jelenleg az R1b, a J-
és, a G-s, és E-s haplocsoportok!
És ez az R1b jelen volt már Sumériában is a J
és G mellett,(zikkuratok),
továbbá R1b-s volt Tutenkhamun fáraó
is,(piramisok)
továbbá ez az R1b-s, J-s, G-s, E-s társaság
építhette a megalitokat Nyugat-Európában,s az R1b-sek a kelta népekben élnek
tovább.
Ezektől már jó 10 ezer éve elkülönült az R1a-s
társaság!
Ők Európa, keleti és középső részeit vették
birtokba, vegyülve az itt őshonos I-sekkel, esetleg N-esekkel.
Ám ha annyira ők voltak a szkíták, azon a területen
ahol mind a mai napig, és most is élnek, miért nincsenek megalitok, és
kurgánok??
És miért fosikálták össze magukat kínjukban, mikor a
kalandozó magyarok rájuk törtek?
Hiszen ők a nagy indoárják, a szkíták
leszármazottai! Ők ötölték ki a lószelídítést, a lovas harcmodort, a kereket!!
Csak mikor ottan hagyták Szkítiját, meg Indiját, s
átkavartak Erjópába hirtelen minden kimént a fejükből, lekopott róluk az
összes dauer, ugye?
Holott még Indiában szorgosan rótták a Védákat
meg az Upanisádokat is :)
Na jó, nem hergelem magam tovább, csak valahogy nem
állom azt a hetyke bajszos szobafestőt, aki sorra irtotta a derék, kazáriai
askenázokat, a világ legnagyobb szellemi tolvajlására alapozódva...
Búcsúzóul még 2videó, az R1b-s rokonságot
alátámasztandó székelyek és kelták között...
Az egyik a Küküllő mentén termett...
A másik Skóciában...
Durva, nemde?
Tutti, hogy ugyannak az ősnótának a variánsai!
Még a szövegben is tetten érhetőek az azonos
motívumok.
De erről majd legközelebb.
Elfáradtam :)